
 PDF下载 ( 833 KB)
PDF下载 ( 833 KB)

腺相关病毒载体在肝纤维化中的应用前景
DOI: 10.12449/JCH260429
-
摘要: 肝纤维化是多种慢性肝病进展为肝硬化和肝癌的关键病理环节,其特征为肝星状细胞活化和细胞外基质过度沉积。目前临床治疗手段有限,而基因治疗作为一种新兴干预策略,正日益受到关注。腺相关病毒(AAV)凭借其低免疫原性、良好的组织靶向性和长期稳定的基因表达能力,已成为理想的基因治疗载体。本文系统综述了AAV载体的结构与功能特点、不同血清型的组织靶向性以及重组AAV的制备与纯化方法,重点阐述了AAV介导的基因过表达、基因沉默、基因编辑和细胞重编程在肝纤维化机制研究与干预中的应用进展,并探讨了其在临床转化中面临的挑战与应对策略。AAV介导的基因治疗技术有望为肝纤维化的精准治疗和机制研究提供新工具和新思路。Abstract: Liver fibrosis is a critical pathological stage in the progression of various chronic liver diseases to liver cirrhosis and liver cancer, and it is characterized by the activation of hepatic stellate cells and excessive deposition of extracellular matrix. There are currently limited clinical treatment methods for liver fibrosis, and gene therapy has attracted increasing attention as a promising intervention strategy. Owing to its advantages of low immunogenicity, strong tissue tropism, and long-term gene expression, adeno-associated virus (AAV) has become an ideal vector for gene therapy. This article systematically reviews the structural and functional features of AAV, the tissue-targeting properties of different serotypes, and the methods for preparation and purification of recombinant AAV and focuses on the advances in the application of AAV-mediated gene overexpression, gene silencing, gene editing, and cellular reprogramming in mechanism research and intervention of liver fibrosis, as well as the challenges in clinical translation and corresponding strategies. AAV-mediated gene therapy is expected to provide novel tools and strategies for the precise treatment and mechanism study of liver fibrosis.
-
Key words:
- Hepatic Fibrosis /
- Dependovirus /
- Genetic Therapy
-
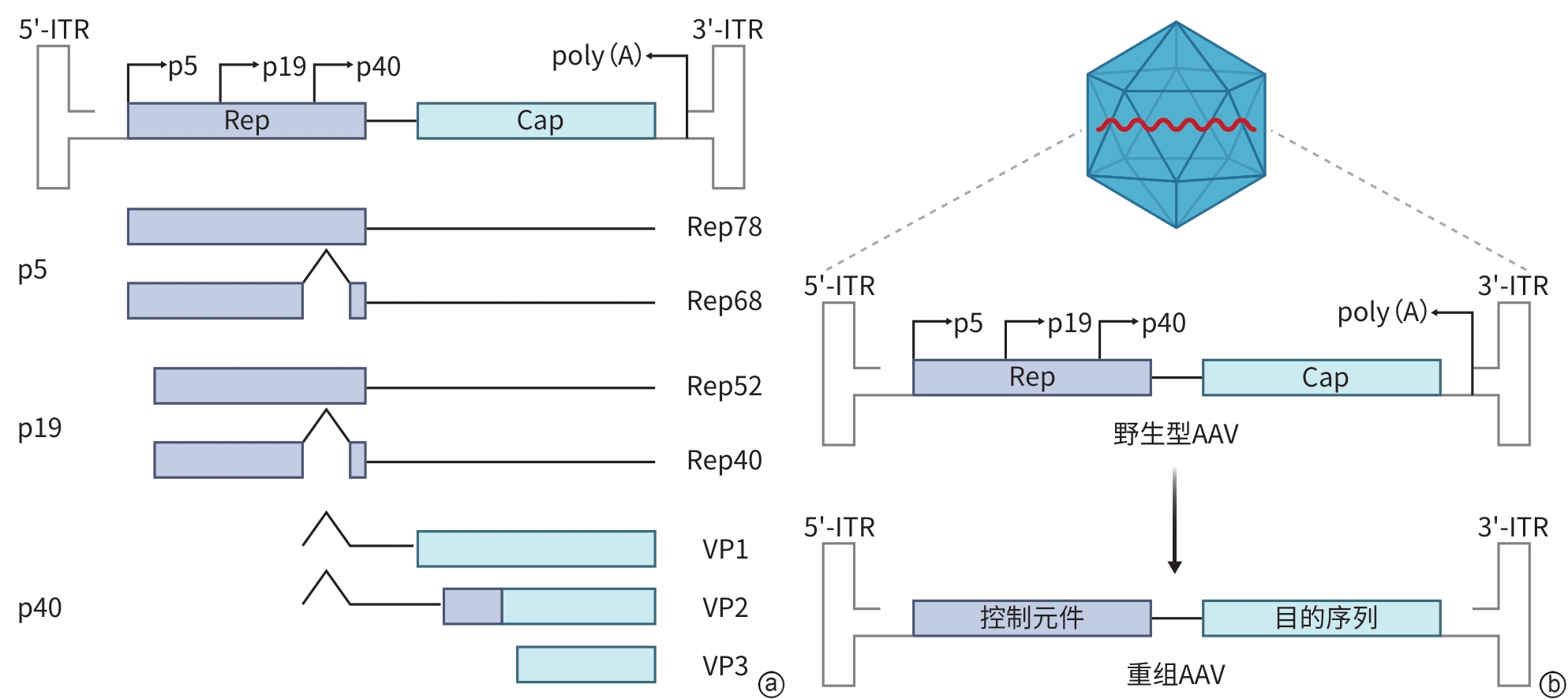
注: a,AAV基因组结构及蛋白表达机制;b,重组AAV载体的构建。ITR,反向末端重复序列;Rep,复制相关蛋白基因;Cap,衣壳蛋白基因;p5、p19,Rep基因上游启动子5、19;p40,Cap基因启动子40;VP1/VP2/VP3,衣壳蛋白1/2/3;poly(A),聚腺苷酸尾;AAV,腺相关病毒。
图 1 AAV的基因组结构及其蛋白表达调控机制
Figure 1. AAV genomic structure and its protein expression regulatory mechanisms
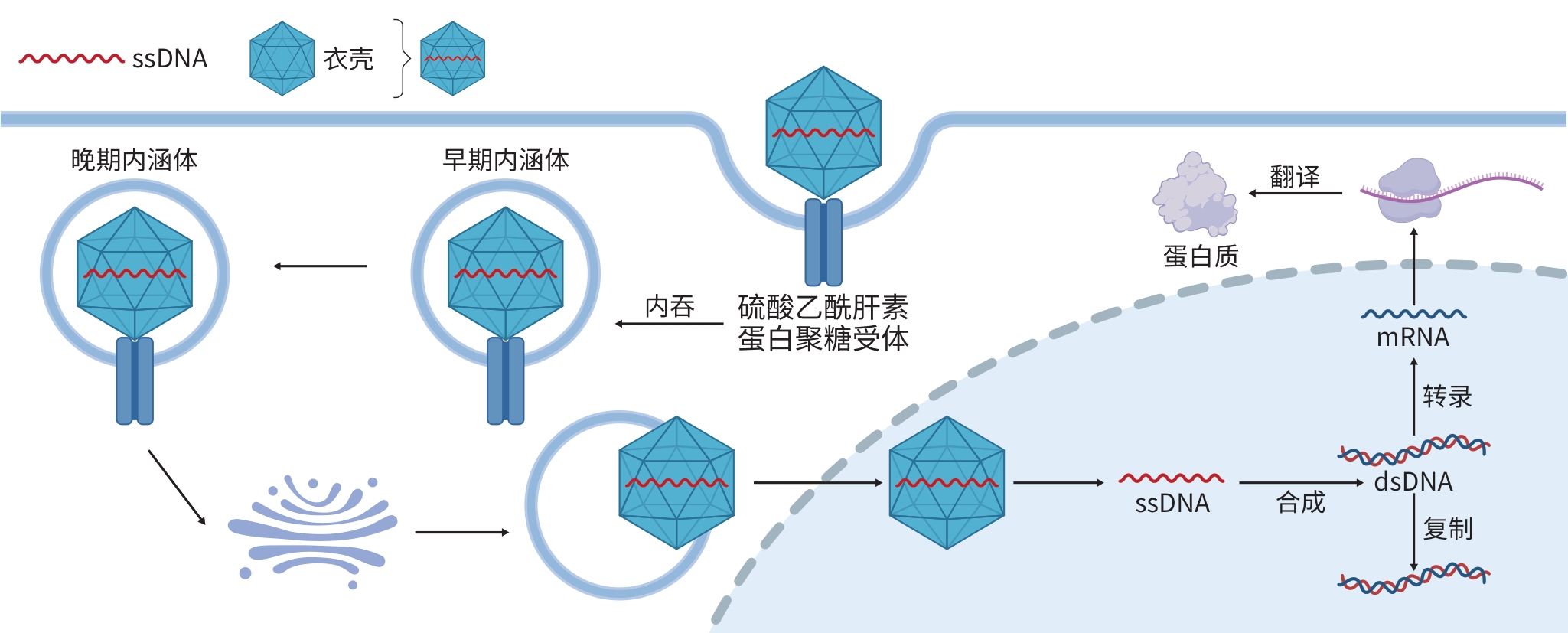
注: ssDNA,单链DNA;dsDNA,双链DNA;mRNA,信使RNA;AAV,腺相关病毒。
图 2 AAV的感染机制
Figure 2. Infection mechanism of AAV
表 1 AAV载体在肝纤维化基因治疗中的核心应用策略与机制
Table 1. Core application strategies and mechanisms of AAV vectors in gene therapy for liver fibrosis
干预策略 作用目的 靶向细胞 作用机制 代表性载体设计 关键疗效指标 基因过
表达纠正代谢紊乱 肝细胞 过表达GNMT,调节单碳代
谢,减少促纤维化因子SAMAAV8-TBG-GNMT ALT/AST下降约60%/55%,
胶原沉积减少约50%[17]清除促炎信号 肝细胞 表达scFv-E06抗体片段,中
和OxPCAAV8-TBG-scFv-E06 阻止单纯脂肪肝向肝纤维
化进展[4]抗氧化应激 肝细胞 过表达Bdh1,激活Nrf2通
路,增强内源性抗氧化能力AAV9-TBG-Bdh1 血清ALT/AST下降,肝纤维
化显著减轻[18]抑制炎症信号 肝细胞 过表达DUSP22,抑制FAK/
MAPK通路AAV8-TBG-Cre 减轻饮食诱导的肝脏炎症
与纤维化[19]调节免疫微环境 肝细胞 过表达HuR,稳定FGF19
mRNA,抑制库普弗细胞M1
极化AAV9-HuR Sirius Red染色总胶原面积
和Collagen Ⅰ表达下降,库
普弗细胞浸润减少[20]基因沉默 抗肝细胞衰老 肝细胞 敲低ATAD3,恢复线粒体功
能,缓解p53介导的衰老AAV8-TBG-shATAD3 衰老细胞减少70%以上,羟
脯氨酸含量下降40%[21]阻断促纤维化轴 肝细胞 敲低FGF9,阻断FGF9/TGF-
β1协同促肝纤维化作用AAV-shFGF9 肝纤维化与HCC发生
减少[22]恢复自噬流 肝细胞 敲低TRIB3,恢复肝细胞自
噬,减少促纤维化外泌体AAV-shTRBI3 减轻BDL/TAA模型肝纤
维化[23]抗HSC活化 HSC HSC特异性表达miR-214,
抑制其增殖与ECM生成AAV8-GFAP-miR-214 有效减轻肝纤维化,未加重
脂肪变性/炎症[24]改善肝窦功能 LSEC细胞 特异性敲低LSEC中ATG5/
RAB27a,增加抑制性EV
释放si-ATG5/RAB27a AAV 肝纤维化面积减少,
α-SMA表达下降[25]基因编辑 永久阻断通路 肝细胞 CRISPR/Cas9直接敲除
TGF-β1, CTGF等核心致病
基因AAV8-CRISPR/SaCas9 肝组织胶原区域减少65%[26] 改善线粒体功能 肝细胞 敲低MCU AAV8-sgMCU 抑制HSC活化与胶原沉积[27] 提升编辑安全性 肝细胞 采用双载体系统限制Cas9
表达,并通过高灵敏度技术
验证其特异性基于AAV8的双载体
系统无脱靶/随机整合;编辑效率
约40%;编辑活性相对
瞬时[28] 下载: 导出CSV
下载: 导出CSV
表 1 (续)
Table 1. (continued)
干预策略 作用目的 靶向细胞 作用机制 代表性载体设计 关键疗效指标 实现转录抑制 肝细胞 递送dCas9-KRAB系统,靶向抑
制致病基因的转录AAV-dCas9-KRAB 靶向抑制Pcsk9,沉默效率>
50%[29]实现转录激活 肝细胞 递送dCas9-VP64系统,靶向激
活保护性基因的转录AAV-dCas9-VP64 成功上调HNF-4α等保护性
基因[29]精准遗传修正 肝细胞 碱基编辑纠正SERPINA1 PiZ
突变,消除Z-AAT聚合物AAV8-eNme2.C-TadA9e AAT聚合物沉积减少,血清
AAT恢复至保护水平[30]靶向治疗
进阶优化递送系统 肝细胞 AAV衣壳工程(如AAV9-GA/
GAST)或与GalNAc偶联的化学
修饰crRNA共递送,以增强肝
脏靶向性、稳定性及编辑效率AAV/GalNAc-crRNA
共递送系统肝脏脱靶DNA水平显著下
降;分次给药可显著提升肝
脏编辑效率[31]体内细胞重编程 肌成纤维
细胞递送肝转录因子,将肌成纤维
细胞直接转化为肝细胞AAV-转录因子组合 逆转肝纤维化,实现组织
再生[32]联合治疗/微环境
重塑肝细胞 沉默c-Met,重塑硬化微环境,
增强干细胞移植疗效AAV-shc-Met+干细胞 干细胞用量减少,胶原沉积
减少[33]注:GNMT,甘氨酸N-甲基转移酶;SAM,S-腺苷甲硫氨酸;TBG,甲状腺素结合球蛋白启动子;ALT,丙氨酸氨基转移酶;AST,天冬氨酸氨基转移酶;scFv-E06,单链可变区抗体片段E06;OxPC,氧化磷脂酰胆碱;Bdh1,β-羟丁酸脱氢酶1;Nrf2,核因子E2相关因子2;DUSP22,双特异性蛋白磷酸酶22;FAK,黏着斑激酶;MAPK,丝裂原激活的蛋白激酶;FGF19,成纤维生长因子19;Sirius Red,天狼星红;Collagen Ⅰ,Ⅰ型胶原;ATAD3,线粒体ATP酶家族蛋白3;p53,抑癌蛋白;FGF9,成纤维生长因子9;TGF-β1,转化生长因子β1;TRIB3,应激蛋白TRIB3;BDL,胆管结扎;TAA,硫代乙酰胺;HSC,肝星状细胞;ECM,细胞外基质;LSEC,肝窦内皮细胞;ATG5,自噬相关基因5;RAB27a,RAS相关蛋白;EV,细胞外囊泡;α-SMA,α-平滑肌肌动蛋白;CTGF,结缔组织生长因子;MCU,线粒体钙单向转运体;Cas9,CRISPR相关蛋白9;Pcsk9,前蛋白转化酶枯草溶菌素9;HNF-4α,肝核因子4α;SERPINA1 PiZ突变,丝氨酸蛋白酶抑制因子家族A成员1 Z等位基因PiZ突变;Z-AAT,Z型α1-抗胰蛋白酶;AAT,α1-抗胰蛋白酶;GalNAc,N-乙酰半乳糖胺;c-Met,细胞间质上皮转换因子;SaCas9,金黄色葡萄球菌Cas9;HCC,肝细胞癌。
下载: 导出CSV
-
[1] MARCELLIN P, KUTALA BK. Liver diseases: A major, neglected global public health problem requiring urgent actions and large-scale screening[J]. Liver Int, 2018, 38( Suppl 1): 2- 6. DOI: 10.1111/liv.13682. [2] KISSELEVA T, BRENNER D. Molecular and cellular mechanisms of liver fibrosis and its regression[J]. Nat Rev Gastroenterol Hepatol, 2021, 18( 3): 151- 166. DOI: 10.1038/s41575-020-00372-7. [3] FRIEDMAN SL, PINZANI M. Hepatic fibrosis 2022: Unmet needs and a blueprint for the future[J]. Hepatology, 2022, 75( 2): 473- 488. DOI: 10.1002/hep.32285. [4] UPCHURCH CM, YEUDALL S, PAVELEC CM, et al. Targeting oxidized phospholipids by AAV-based gene therapy in mice with established hepatic steatosis prevents progression to fibrosis[J]. Sci Adv, 2022, 8( 28): eabn0050. DOI: 10.1126/sciadv.abn0050. [5] MOSCOSO CG, STEER CJ. Liver targeted gene therapy: Insights into emerging therapies[J]. Drug Discov Today Technol, 2019, 34: 9- 19. DOI: 10.1016/j.DDTec.2020.11.001. [6] ZHAO Z, LIN CY, CHENG K. siRNA- and miRNA-based therapeutics for liver fibrosis[J]. Transl Res, 2019, 214: 17- 29. DOI: 10.1016/j.trsl.2019.07.007. [7] YANG Z, YAO YZ, CHEN X, et al. Cross-species tropism of AAV.CPP.16 in the respiratory tract and its gene therapies against pulmonary fibrosis and viral infection[J]. Cell Rep Med, 2025, 6( 6): 102144. DOI: 10.1016/j.xcrm.2025.102144. [8] XIAO ZH, WANG YJ, CHEN YY, et al. Exosomes derived from TREM-2 knocked-out macrophages alleviated renal fibrosis via HSPa1b/AKT pathway[J]. Am J Physiol Renal Physiol, 2025, 328( 1): F131- F151. DOI: 10.1152/ajprenal.00219.2024. [9] YUAN SJ, YANG SJ, GUO YJ, et al. GLIPR2 regulates EndoMT and cardiac fibrosis after AMI via PDGFRL/AKT/mTOR signaling pathway[J]. Life Sci, 2025, 378: 123816. DOI: 10.1016/j.lfs.2025.123816. [10] MEYER NL, CHAPMAN MS. Adeno-associated virus(AAV) cell entry: Structural insights[J]. Trends Microbiol, 2022, 30( 5): 432- 451. DOI: 10.1016/j.tim.2021.09.005. [11] SANTIAGO-ORTIZ JL, SCHAFFER DV. Adeno-associated virus(AAV) vectors in cancer gene therapy[J]. J Control Release, 2016, 240: 287- 301. DOI: 10.1016/j.jconrel.2016.01.001. [12] PUPO A, FERNÁNDEZ A, LOW SH, et al. AAV vectors: The Rubik's cube of human gene therapy[J]. Mol Ther, 2022, 30( 12): 3515- 3541. DOI: 10.1016/j.ymthe.2022.09.015. [13] WÖRNER TP, BENNETT A, HABKA S, et al. Adeno-associated virus capsid assembly is divergent and stochastic[J]. Nat Commun, 2021, 12( 1): 1642. DOI: 10.1038/s41467-021-21935-5. [14] KUZMIN DA, SHUTOVA MV, JOHNSTON NR, et al. The clinical landscape for AAV gene therapies[J]. Nat Rev Drug Discov, 2021, 20( 3): 173- 174. DOI: 10.1038/d41573-021-00017-7. [15] MATHIESEN SN, LOCK JL, SCHODERBOECK L, et al. CNS transduction benefits of AAV-PHP.eB over AAV9 are dependent on administration route and mouse strain[J]. Mol Ther Methods Clin Dev, 2020, 19: 447- 458. DOI: 10.1016/j.omtm.2020.10.011. [16] KISH WS, LIGHTHOLDER J, ZEKOVIĆ T, et al. Removal of empty capsids from high-dose adeno-associated virus 9 gene therapies[J]. Biotechnol Bioeng, 2024, 121( 8): 2500- 2523. DOI: 10.1002/bit.28737. [17] FANG CC, WU CF, LIAO YJ, et al. AAV serotype 8-mediated liver specific GNMT expression delays progression of hepatocellular carcinoma and prevents carbon tetrachloride-induced liver damage[J]. Sci Rep, 2018, 8( 1): 13802. DOI: 10.1038/s41598-018-30800-3. [18] XU BT, TENG FY, WU Q, et al. Bdh1 overexpression ameliorates hepatic injury by activation of Nrf2 in a MAFLD mouse model[J]. Cell Death Discov, 2022, 8( 1): 49. DOI: 10.1038/s41420-022-00840-w. [19] GE CX, TAN J, DAI XL, et al. Hepatocyte phosphatase DUSP22 mitigates NASH-HCC progression by targeting FAK[J]. Nat Commun, 2022, 13( 1): 5945. DOI: 10.1038/s41467-022-33493-5. [20] MO XQ, ZHOU SJ, ZHOU XG, et al. HuR enhances the stability of FGF19 mRNA to suppress Kupffer cell activation and mitigate inflammation and fibrosis in non-alcoholic fatty liver disease[J]. Gen Physiol Biophys, 2025, 44( 5): 349- 361. DOI: 10.4149/gpb_2025023. [21] CHEN LT, LI YC, PATEL RN, et al. AAV8-mediated silencing of Atad3 prevents the progression from simple steatosis to MASH in mice by reduced IL6 secretion[J]. Biochim Biophys Acta Mol Basis Dis, 2025, 1871( 5): 167792. DOI: 10.1016/j.bbadis.2025.167792. [22] ZHANG L, ZHANG Q, TENG D, et al. FGF9 recruits β-catenin to increase hepatic ECM synthesis and promote NASH-driven HCC[J]. Adv Sci, 2023, 10( 28): 2301166. DOI: 10.1002/advs.202301166. [23] ZHANG XW, ZHOU JC, PENG D, et al. Disrupting the TRIB3-SQSTM1 interaction reduces liver fibrosis by restoring autophagy and suppressing exosome-mediated HSC activation[J]. Autophagy, 2020, 16( 5): 782- 796. DOI: 10.1080/15548627.2019.1635383. [24] ZHAO FQ, NIU X, SONG G, et al. Hepatic stellate cell-specific miR-214 expression alleviates liver fibrosis without boosting steatosis and inflammation[J]. J Transl Med, 2025, 23( 1): 810. DOI: 10.1186/s12967-025-06880-x. [25] CHEN TT, ZHANG Y, ZHANG YJ, et al. Autophagic degradation of MVBs in LSECs promotes Aldosterone induced-HSCs activation[J]. Hepatol Int, 2024, 18( 1): 273- 288. DOI: 10.1007/s12072-023-10559-0. [26] YAN K, FENG JP, LIU X, et al. Inhibition of hepatitis B virus by AAV8-derived CRISPR/SaCas9 expressed from liver-specific promoters[J]. Front Microbiol, 2021, 12: 665184. DOI: 10.3389/fmicb.2021.665184. [27] LI SY, CHEN FY, LIU M, et al. Knockdown of hepatic mitochondrial calcium uniporter mitigates MASH and fibrosis in mice[J]. Cell Biosci, 2024, 14( 1): 135. DOI: 10.1186/s13578-024-01315-4. [28] SINGH K, FRONZA R, EVENS H, et al. Comprehensive analysis of off-target and on-target effects resulting from liver-directed CRISPR-Cas9-mediated gene targeting with AAV vectors[J]. Mol Ther Methods Clin Dev, 2024, 32( 4): 101365. DOI: 10.1016/j.omtm.2024.101365. [29] WANG D, ZHANG F, GAO GP. CRISPR-based therapeutic genome editing: Strategies and in vivo delivery by AAV vectors[J]. Cell, 2020, 181( 1): 136- 150. DOI: 10.1016/j.cell.2020.03.023. [30] GAO J, BAMIDELE N, PIRES-FERREIRA D, et al. A compact base editor rescues AATD-associated liver and lung disease in mouse models[J]. Mol Ther, 2025, 33( 11): 5817- 5828. DOI: 10.1016/j.ymthe.2025.08.040. [31] ZHANG H, KELLY K, LEE J, et al. Self-delivering, chemically modified CRISPR RNAs for AAV co-delivery and genome editing in vivo[J]. Nucleic Acids Res, 2024, 52( 2): 977- 997. DOI: 10.1093/nar/gkad1125. [32] REZVANI M, ESPAÑOL-SUÑER R, MALATO Y, et al. In vivo hepatic reprogramming of myofibroblasts with AAV vectors as a therapeutic strategy for liver fibrosis[J]. Cell Stem Cell, 2016, 18( 6): 809- 816. DOI: 10.1016/j.stem.2016.05.005. [33] ZHANG HY, CHEN QY, HU DY, et al. Manipulating HGF signaling reshapes the cirrhotic liver niche and fills a therapeutic gap in regeneration mediated by transplanted stem cells[J]. Exp Cell Res, 2024, 434( 1): 113867. DOI: 10.1016/j.yexcr.2023.113867. [34] HE WB, SUN ZC, TONG G, et al. FUNDC1 alleviates doxorubicin-induced cardiotoxicity by restoring mitochondrial-endoplasmic reticulum contacts and blocked autophagic flux[J]. Theranostics, 2024, 14( 9): 3719- 3738. DOI: 10.7150/thno.92771. [35] BING SJ, SEIRUP M, HOANG TT, et al. Rational immunosilencing of a promiscuous T-cell epitope in the capsid of an adeno-associated virus[J]. Nat Biomed Eng, 2024, 8( 2): 193- 200. DOI: 10.1038/s41551-023-01129-8. [36] KUMAR SRP, BISWAS M, CAO D, et al. TLR9-independent CD8+ T cell responses in hepatic AAV gene transfer through IL-1R1-MyD88 signaling[J]. Mol Ther, 2024, 32( 2): 325- 339. DOI: 10.1016/j.ymthe.2023.11.029. [37] WANG JH, GESSLER DJ, ZHAN W, et al. Adeno-associated virus as a delivery vector for gene therapy of human diseases[J]. Signal Transduct Target Ther, 2024, 9( 1): 78. DOI: 10.1038/s41392-024-01780-w. [38] SUORANTA T, LAHAM-KARAM N, YLÄ-HERTTUALA S. Strategies to improve safety profile of AAV vectors[J]. Front Mol Med, 2022, 2: 1054069. DOI: 10.3389/fmmed.2022.1054069. -

 本文二维码
本文二维码
计量
- 文章访问数: 217
- HTML全文浏览量: 124
- PDF下载量: 66
- 被引次数: 0

